
Первый уровень дифференциации пола в развитии
Обновлено: 27.04.2024
Процесс половой дифференциации начинается с оплодотворения яйцеклетки и проходит ряд стадий, каждая из которых имеет свои задачи, результаты которых становятся необратимыми. Генетический пол определяет, обусловленным строением половой железы (яичка или яичника). Возникновение яичек или яичников обусловливает гаметный пол. Возникает гормональный пол, который у зародыша определяет дифференциацию внутренних репродуктивных органов и наружных гениталий, а также особых нервных механизмов ("половых центров"), которые в будущем регулируют поведение человека. В период полового созревания выделяется большое количество гормонов, которые окончательно определяют биологические различия по полу. У мальчиков уровень тестостерона увеличивается в 18 раз, а у девочек уровень эстрадиола - в 8 раз. При отсутствии или недостатке в этот критический период зародышевых андрогенов половая дифференциация автоматически, независимо от хромосомного пола, происходит по женскому типу.. Женский псевдо гермафродитизм выражается в увеличение клитора. Такая девочка при рождении записывается и воспитывается как мальчик до тех пор, пока не начинается период полового созревания и не появляются грудные железы и менструации. Мужской псевдо гермафродитизм связан с недостатком мужских половых гормонов в период развития мальчиков. Вследствие этого мальчики приобретают некоторые морфологические и поведенческие черты, свойственные женщинам.
Гипоталамус, в котором расположены половые центры, не только дифференцируется под влиянием зародышевых гормонов, но и сам является психоэндокринным органом; его пренатальная программа, ориентированная на мужское и женское поведение, определяет характер его реакции на половые гормоны пубертатного периода, а эта реакция, в свою очередь, вызывает соответствующее полодиморфическое поведение.
В период полового созревания выделяется большое количество гормонов, окончательно определяющих биологические различия по полу. За этот период у мальчиков уровень тестостерона увеличивается в 18 раз, а у девочек уровень эстрадиола — в 8 раз.
При отсутствии или недостатке в соответствующий критический период зародышевых андрогенов половая дифференциация автоматически, независимо от хромосомного пола, происходит по женскому типу. Примером может послужить развитие ребенка в тех случаях, когда в силу патологического влияния экологии (интоксикация, радиация) не формируются половые железы (состояние агонадизма).С другой стороны, если мать в период беременности принимает препараты, стимулирующие появление мужского гормона (тестостерона), то женский эмбрион может «дефеминизироваться», что впоследствии проявится в маскулинизации женского поведения. Такие девочки предпочитают общество мальчиков и игры, свойственные мальчикам, они самоуверенны и независимы, т. е. определяются как сорванцы. Все это доказывает, что андрогены играют существенно большую роль для внутриутробной дифференцировки полов, чем эстрогены.

Различия в быстроте развития мужского и женского организмов видны уже на стадии эмбриона. У девочек развитие скелета идет быстрее. После рождения они на 1-2 недели опережают мальчиков в формировании костей. В то же время по длине и весу мальчики при рождении больше девочек на 2-3%. (Ильин, психофизиология)
Дифференциация пола – это процесс формирования морфофизиологических и поведенческих различий между полами в онтогенезе. Она представляет собой цепь закономерно сменяющих друг друга этапов, причем каждый последующий этап основывается на предыдущем.
Несмотря на разнообразие вариантов у разных животных, принципиальная схема дифференцировки весьма консервативна и представляет определенную последовательность событий. Необходимо отметить, что развитие в направлении женского организма – это «нормальная судьба» зиготы. Для развития в мужском направлении необходимы особые «переключатели», подавляющие самодифференциацию в женском направлении. В онтогенезе млекопитающих можно выделить 3 основных этапа дифференцировки.
На 1-м этапе на основе детерминации пола происходит дифференциация гонад.
На 2-м этапе дифференцированные гонады выделяют гормоны, которые активируют гены, обусловливающие половую дифференциацию фенотипа и мозга.
На 3-м этапе вследствие дифференциации мозга и процессов социализации формируется половое поведение.
Дифференциация гонад. У эмбриона млекопитающих происходит закладка бисексуальной системы эмбриональных гонад и двух пар половых протоков – мюллеровых и вольфовых каналов. Под действием гена SRY эмбриональные гонады преобразуются в семенники, а в случае отсутствия этого гена – в яичники.
После дифференцировки эмбриональных гонад в семенники они начинают выделять гормоны – тестостерон и антимюллеровский гормон (АМН). АМН обусловливает дегенерацию мюллеровых каналов. Тестостерон обусловливает преобразование вольфовых каналов в семявыносящие протоки и придатки яичек; его можно считать вторым «главным переключателем», так как он является важнейшим фактором маскулинизации. Кастрированные на ранних стадиях зародыши развиваются по женскому типу, независимо от кариотипа.
При дифференцировке эмбриональных гонад в яичники не происходит выделения тестостерона и АМН. Это сопровождается запрограммированным преобразованием мюллеровых каналов в женскую проводящую систему, состоящую из матки и яйцеводов. Воль-фовы каналы дегенерируют.
Дифференциация гамет. Дифференциация гамет – процесс преобразований первичных половых клеток (ППК, или гоноцитов) в мужские (сперматогонии) или женские (оогонии) половые клетки. Этот процесс независим от дифференциации гонад и определяется половым кариотипом.
Давно доказано внегонадное происхождение первичных половых клеток позвоночных. У млекопитающих они образуются в эктодерме желточного мешка, откуда мигрируют вначале к задней части зародыша, а затем – в область закладки гонад. Во время своего «путешествия» ППК делятся. После дифференцировки гонад ППК также дифференцируются и превращаются либо в сперматогонии, либо в оогонии, в зависимости от пола гонад.
Процессы дифференциации гамет и гонад являются примером такого явления как автономность развития частей единой структуры. Это типично для процессов онтогенеза: другим примером может служить независимость формирования индуктора и компетентной ткани. Целостность морфогенеза достигается синхронизацией событий. В случае десинхронизации обычно наблюдаются различные пороки развития.
Фенотипическая половая дифференциация. Развитие вторичных половых признаков у млекопитающих контролируется многими генами. Одним из основных является ген Tfm, локализованный на Х-хромосоме. Это ген рецептора андрогенов. Его регуляторные белки, связываясь с тестостероном, активируют гены, необходимые для дифференцировки по мужскому типу.
Локализация гена Tfm на Х-хромосоме показывает, что клеточные рецепторы тестостерона должны быть как на XX-, так и на ХY-кариотипах. Поэтому введение тестостерона в зародыши XX вызывает развитие вторичных половых признаков самца. Однако отсутствие гормона АМН у таких зародышей приводит к развитию как женских, так и мужских половых путей и формированию гермафродитизма.
В результате половой дифференциации формируются:
Первичные половые признаки – морфофизиологические характеристики половой системы у разных полов.
Вторичные половые признаки – морфофизиологические характеристики фенотипов разных полов, не относящихся к половой системы.
Степень различия между полами по вторичным половым признакам получила название половой диморфизм.
Гермафродитизм – это направление в процессах дифференциации пола, приводящее к формированию организмов с признаками обоих полов.
Половая дифференциация мозга и поведения. Главным фактором маскулинизации мозга и опосредуемого им поведения является также тестостерон. Опыты на животных показали, что самцы, кастрированные на ранней стадии, демонстрируют поведение, характерное для самок.
Для маскулинизации мозга тестостерон должен воздействовать в особые чувствительные критические периоды онтогенеза, причем его отсутствие в это время не может быть компенсировано в дальнейшем. Именно своевременное воздействие этого гормона закладывает базовые основы полового поведения.
Если формы полового поведения у беспозвоночных обычно стереотипны и жестко детерминированы, то у высших позвоночных они демонстрируют широкий диапазон индивидуальной изменчивости под влиянием сообщества и научения. Многочисленные работы этологов показали роль импринтинга, изоляции, общения с матерью и сородичами в последующем формировании полового поведения у птиц и млекопитающих. У млекопитающих особое значение имеет контакт с матерью. Однако, даже выращенные с матерью, но в изоляции от сверстников самцы, часто оказываются неспособными к спариванию, к установлению коммуникаций.
Половая социализация – это процесс формирования моделей полового поведения в ходе постнатального развития.
Для социальных животных возможность спаривания самым тесным образом связана с их положением в группе, поэтому процесс социализации приобретает важнейшее значение. На протяжении постнатального онтогенеза половая социализация может представлять особый, длительный и многоэтапный процесс, где каждый последующий этап зависит от предыдущего и где имеются свои критические периоды.
Необычайно широк репертуар полового поведения у человека вследствие влияния разнообразных факторов культуры.
В процессах дифференциации, как и детерминации, особое значение имеет понятие критического периода. Каждый этап дифференциации пола происходит только в определенный период развития организма. Если такой критический период пропущен, т. е. в необходимое время запускающие сигналы отсутствовали, то последствия обычно необратимы. Поскольку разные этапы дифференцировки запускаются различными пусковыми механизмами и в разное время, то в онтогенезах наблюдаются многочисленные варианты отклонений. Особенно часто такие отклонения встречаются при формировании половых признаков и полового поведения.
Принадлежность организмов к тому или иному полу часто является результатом сложного взаимодействия генетических, экологических, физиологических, а иногда и психологических факторов. Однако решающее значение имеет тот «выбор», который осуществляется при детерминации.
Детерминация пола – исходное направление развития организма в сторону мужского или женского пола вследствие определенных факторов.
У высших организмов генетическая детерминация обычно выражается мужским или женским кариотипом зиготы, образующимся в момент оплодотворения. Наблюдается несколько вариантов генетической детерминации пола, но наиболее общая тенденция – это различные сочетания половых хромосом у разных полов.
Варианты половых кариотипов в природе рассматривались нами ранее. Однако не сам половой кариотип является непосредственным фактором детерминации пола. Этим фактором служит стартовый сигнал, воспринимаемый «ключевым» геном. Одинаковые половые кариотипы могут участвовать в разных механизмах детерминации. Таких механизмов в природе встречается множество. У высших животных процессы формирования пола проходят более сложно, но единообразнее. Рассмотрим некоторые варианты детерминации пола.
У дрозофилы контролирующим сигналом является соотношение числа Х-хромосом и гаплоидных наборов аутосом. Соотношение 1 дает самок, а 0,5 – самцов. Анеуплоидия по половым хромосомам, сдвигающая это соотношение, дает либо интерсексов, либо бесплодных особей с гипертрофированными половыми признаками (сверхсамка и сверхсамец). Это открытие, сделанное американским генетиком К. Бриджесом в 1921 г., легло в основу балансовой теории детерминации пола. Ключевым геном, «улавливающим» баланс хромосом кариотипа дрозофилы(по соотношению белковых продуктов определенных генов), является ген Sxl (Sex lethal). В зависимости от баланса хромосом ген Sxl экспрессируется по-разному, и сложные каскады других регуляторных генов расходятся в направлении мужского или женского пола.
Широко представлена в природе, в частности у многих рептилий, и так называемая экологическая детерминация, когда пол определяется внешними условиями (температурой, продолжительностью светового дня). При этом создается впечатление независимости детерминации от генетических характеристик. Но современная биология развития рассматривает влияние внешней среды как сигнал, «включающий» или «выключающий» ключевой ген. На роль такого стартового гена-переключателя, запускающего генетические механизмы определения пола у яйцекладущих форм, претендует гипотетический фактор Testis Determining Factor (TDF). Вероятно, он кодирует белки, регулирующие экспрессию важнейших генов, участвующих в процессе детерминации пола.
У млекопитающих, имеющих одинаковые с дрозофилой половые кариотипы, мы видим другой механизм детерминации пола, в котором основное значение имеет наличие Y-хромосомы. Она направляет развитие в сторону мужского пола при любом количестве Х-хромосом.
Эти наблюдения показывают, что в процессах детерминации пола млекопитающих задействованы не только гены Y-хромосомы, но и гены Х-хромосомы и аутосом. Существует, например, белковый фактор SF-1 – регулятор генов для всех ферментов синтеза стероидов, к которым относятся половые гормоны. Дефект гена SF-1 может привести к отсутствию дифференцированных гонад у обоих полов независимо от кариотипа.
Таким образом, при детерминации пола особенно трудно провести четкие временные границы. Можно добавить, что до сих пор во многом не ясно соподчинение генетических механизмов детерминации.
Половая дифференциация человека. Мужской пол. Женский пол. Генетический пол. Аутосомы. Половой диморфизм.
При фертилизации, в зиготе, находящейся в репродуктивном тракте женского организма, на основе генетической программы первоначально формируется хромосомный, или генетический, пол человека. Затем в ходе развития эмбриона формируется гонадный пол, когда недифференцированные гонады развиваются либо в семенники, либо в яичники. В зависимости от гонадного пола у плода происходит дифференциация мужских или женских внутренних и наружных гениталий (формируется фенотипиче-ский пол). В целом указанные превращения называется половой дифференциацией человека (рис. 16.2).
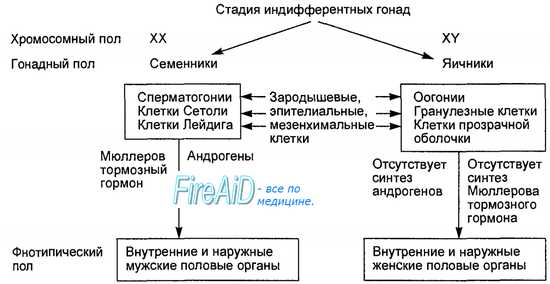
Рис. 16.2. Половая дифференциация человека. Эмбрион имеет мужской (XY) и женский (XX) хромосомный пол. Хромосомный пол сменяется развитием индифферентных гонад в яичники или семенники. В результате формируется гонадный пол. Семенники секретируют тестостерон, который обусловливает развитие мужского фенотипического пола, а отсутствие секреции тестостерона обусловливает развитие женского фенотипического пола.
Генетический пол
Половые хромосомы (X и Y) являются единственными из 23 хромосом, которые определяют пол организма. Половые клетки, продуцируемые женским организмом, содержат Х-хромосому, тогда как сперматозоиды — X-или Y-хромосому. Остальные 22 хромосомы предопределяют характеристики организма, не связанные с полом, и поэтому называются аутосомами.
Во время фертилизации образуется зигота, содержащая 23 пары хромосом. В зиготе, из которой будет развиваться плод женского пола, содержится две Х-хромосомы, т. е. по одной из каждой женской и мужской гаметы. В зиготе, из которой будет развиваться плод мужского пола, содержится Х- и Y-хромосомы. Развитие гонад и гениталий в мужском организме обусловлено Y-хромосомой. При отсутствии в генах эмбриона Y-хромосомы у эмбриона развиваются яичники. Y-хромосома ответственна за начальный половой диморфизм, т. е. две различные половые формы, мужская и женская, одного и того же индивидуума.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Гонадный пол. Индифферентная стадия развития эмбриона. Особенности формирования гонадного пола у плода.
Примерно до 6—7-й недели внутриутробной жизни эмбриона его гонады развиваются идентично как у мужчин, так и у женщин. Это так называемая индифферентная стадия развития эмбриона, когда половая система обоих полов развивается по одной генетической программе. В период формирования гонадного пола у плода развиваются мужской (вольфов) и женский (мюллеров,) протоки. Первоначально развитие этих протоков начинается унипотенциально, т. е. независимо от будущего пола, и только один из них может развиваться в половой тракт, связанный с конкретным генетическим полом плода. В частности, вольфов проток превращается в структуры мужского полового тракта, а мюллеров — женского (табл. 16.1). Закладка индифферентных гонад происходит у эмбриона в виде гонадных валиков. В эпителии гонадных валиков обособляются крупные первичные половые клетки, которые в дальнейшем дают начало развитию половых клеток. Первичные половые клетки дифференцируются на 5-й день после фертилизации. По прошествии 3 нед они определяются среди клеток внезародышевой энтодермы в стенке желточного мешка (часть первичной кишки, вынесенная за пределы зародыша). От поверхности внутрь половых валиков прорастают тяжи, состоящие из первичных половых клеток фолликулярного эпителия, который выполняет для первичных половых клеток трофическую роль. В течение 3 нед жизни эмбриона в половых валиках эти клеточные элементы развивается в ткань, которая будет основой первичных гонад. Первичные половые клетки из вольфова тела мигрируют в эту ткань, в которой из них будут образовываться половые клетки или гаметы. В индифферентный период развития первичных гонад в их стро-мальной ткани содержится два типа клеток. Один тип клеток на гонадной стадии дифференциации пола развивается в гранулезные клетки яичников или в клетки Сертоли семенных трубочек в яичках. Второй тип клеток на стадии гонад дифференцируются в клетки прозрачной оболочки (theca pellucida) в яичниках или в клетки Лейдига в семенниках.
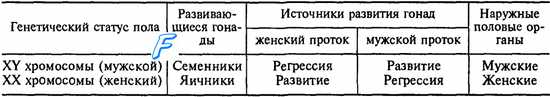
Таблица 16.1. Особенности формирования гонадного пола у плода
У эмбриона мужского пола на 6—7-й неделе эмбриональной жизни после миграции первичных половых клеток в первичные гонады в присутствии Y-хромосомы, содержащей SRY-ген, происходит дифференциация клеток Сертоли. В процессе дифференциации клетки Сертоли располагаются вокруг первичных половых клеток, в результате начинается развитие трубочек семенников в первичных гонадах. Дифференциация мезенхи-мальных (стромальных) клеток гонад в интерстициальные клетки Лейдига, которые в последующем будут секретировать мужской половой гормон тестостерон, начинается с 8—9-й недели и заканчивается на 10-й неделе развития плода.
У женского эмбриона дифференцирование первичных гонад в яичники начинается с 9-й недели, когда активируются Х-хромосомы. Их активация приводит к пролиферации первичных половых клеток, деление которых осуществляется путем митоза. К моменту рождения плода женского пола в яичниках содержатся первичные ооциты, которые образуются в период развития плода из первичных половых клеток путем мейотического деления. Однако уже в периоде внутриутробного развития мейоз первичных половых клеток блокируется (стадия поздней профазы) после образования первичного ооцита. В первичном ооците мейоз на стадии поздней профазы вновь возобновится спустя много лет в одну из будущих овуляций, под влиянием гонадотропинов гипофиза и гормонов половых желез, в периоде пубертатного развития женского организма.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: